


 Minsoy
Minsoy ×
×{kind=link}
{kind=link}
{kind=link}
{kind=link}
优良品系中品03-5373系谱的遗传解析及抗大豆胞囊线虫病相关标记鉴定
引用本文
张姗姗, 李英慧, 李金英, 邱丽娟. 优良品系中品03-5373系谱的遗传解析及抗大豆胞囊线虫病相关标记鉴定. 作物学报, 2013, 39(10): 1746-1753
[ZHANG Shan-Shan, LI Ying-Hui, LI Jin-Ying, QIU Li-Juan. Genetic Dissection of Elite Line Zhongpin 03-5373 Pedigree and Identification of Candidate Markers Related to Resistance to Soybean Cyst Nematode. Acta Agronomica Sinica, 2013, 39(10): 1746-1753]
Permissions
[ZHANG Shan-Shan, LI Ying-Hui, LI Jin-Ying, QIU Li-Juan. Genetic Dissection of Elite Line Zhongpin 03-5373 Pedigree and Identification of Candidate Markers Related to Resistance to Soybean Cyst Nematode. Acta Agronomica Sinica, 2013, 39(10): 1746-1753]
Copyright©2013, Editorial office of Acta Agronomica Sinica
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
优良品系中品03-5373系谱的遗传解析及抗大豆胞囊线虫病相关标记鉴定
摘要
方法
中品03-5373是高抗大豆胞囊线虫(soybean cyst nematode, SCN) 3号生理小种的优良大豆新种质, 可追溯到10个祖先亲本, 其中包括灰皮支黑豆、Peking和PI437654等国内外SCN主要抗源。本研究利用152个SSR标记对中品03-5373及其亲本进行鉴定, 共发现等位变异437个, 每个标记的等位变异范围为2~5个, 平均为2.9个。亲缘关系分析表明11份材料间的遗传一致度变化范围为0.2430~0.8224, 平均为0.458, 4个SSR标记(Satt152、Satt179、Barcsoyssr_18_107和Satt196)形成的单倍型可以将11份材料区别开来。系谱追踪阐明了育种对基因组组成变化的作用, 发现中品03-5373亲本中以灰皮支黑豆贡献的等位变异最多(39个), PI437654次之(6个)。通过系谱追踪筛选到与SCN 3抗性相关的候选标记20个, 为进一步克隆抗病基因和选择有效的标记组合进行分子育种提供了依据。
Method
关键词:
大豆; 胞囊线虫病; 系谱分析; 分子标记
Genetic Dissection of Elite Line Zhongpin 03-5373 Pedigree and Identification of Candidate Markers Related to Resistance to Soybean Cyst Nematode
Abstract
Zhongpin 03-5373 with resistance to SCN3 is an elite soybean line, which is traced back to 10 parental lines. Three elite resistant sources (Huipizhiheidou, Peking, and PI437654), diversely used in the soybean cyst nematode resistant breeding, were included in the pedigree of Zhongpin 03-5373. In this study, 152 molecular markers were selected to detect polymorphism among the 11 materials. A total of 437 alleles were identified with average of 2.9, ranging from two to five alleles on each locus. The average of Nei’s genetic identity among pairwise cultivars were 0.458, the haplotypes formed by four SSR markers (Satt152, Satt179, Barcsoyssr_18_107, and Satt196) could distinguish 11 cultivars in this study. Pedigree tracing elucidated genomic variation in breeding. IBD analysis showed that Huipizhiheidou contributed the most of specific alleles (39) to Zhongpin 03-5373 among three resistant genetic resources (Huipizhiheidou, Peking, and PI437654), followed by PI437654 (6). In addition, 20 markers identified were related to the resistance to SCN 3. These results supplied the information for further cloning resistant gene and resistant breeding by marker assistant selection.
Keyword:
Soybean; Soybean cyst nematode; Pedigree analysis; Molecular marker
引言
大豆胞囊线虫(soybean cyst nematode, SCN)导致世界大豆产量严重损失, 在我国大豆产区发生的面积也在逐年增加, 并有蔓延的趋势[1]。目前我国发现1、2、3、4、5、6、7、9和14号生理小种, 其中3号生理小种是我国东北大豆主产区的优势小种。选育和种植抗性品种是最有效的控制大豆胞囊线虫病害的方法[2,3]。
经典遗传学分析表明, 大豆中有4个抗性位点控制抗大豆胞囊线虫基因。自1994年以来, 已定位到与SCN抗性相关的位点或标记150多个[4]。这些位点或标记皆在双亲的分离群体中鉴定的, 易受双亲遗传背景限制, 定位到的标记普遍应用性有待验证, 只能用于部分品种的杂交后代实验的分子标记辅助选择(marker-assisted selection, MAS), 故利用遗传背景比较广泛的材料筛选与大豆胞囊线虫病抗性相关的分子标记具有重要的意义。
“血缘一致性”或者“同源一致性”(identity by descent, IBD)是鉴定多世代、多杂交组合的系谱材料中同重要农艺性状相关的优异等位变异的统计方法[5]。该方法既充分利用了连锁分析(linkage analysis)所提供的重组信息又考虑了关联分析(association analysis)所提供的连锁不平衡(linkage disequilibrium, LD)信息, 可以分析世代延续过程中基因与其周围标记的重组情况, 在分子标记水平上将品种、育种选择和后代连接起来, 鉴定选育过程中受到选择的位点和区间。该方法曾应用到大麦[6]和大豆[7,8]的重要性状分子标记研究中, 但在抗病基因标记研究中尚未见报道。
中品03-5373是用黄种皮抗SCN4号生理小种的晋1265与引自美国的抗多个SCN小种的Hartwig杂交选育出的既高抗大豆胞囊线虫4号生理小种[9]也对SCN3免疫的优良大豆新种质, 其亲本中有多个国内外大豆胞囊线虫主要优异抗源。本研究利用SSR、EST-SSR、InDel等共显性分子标记分析11份中品03-5373及祖先亲本系谱, 旨在解析中品03-5373和其亲本之间的遗传关系, 挖掘与胞囊线虫3号生理小种抗性相关的标记, 为我国大豆抗胞囊线虫病新品种培育及抗大豆胞囊线虫病基因的挖掘奠定基础。
1 材料与方法
1.1 试验材料
中品03-5373的系谱可追踪到11个亲本(图1), 包括灰皮支黑豆、Hartwig、Peking、PI437654、Forrest、Lee、晋1265、Hill、Dyer和Bragg。灰皮支黑豆、Peking和PI437654等3个祖先亲本是中国和美国广泛用于抗病基因发掘和品种选育的重要抗源, 其中灰皮支黑豆高抗大豆胞囊线虫1、3、4、5号生理小种[10], Peking是美国和中国现在所育成抗大豆胞囊线虫品种中抗病基因的最主要来源, 抗1、3和5号生理小种, PI437654抗大豆胞囊线虫1~5号生理小种。
该系谱中一系列优良的抗病品种抗谱比较广, 其中Dyer高抗1号和3号生理小种, 但产量较低[11]。Forrest是以Dyer为抗病亲本育成的品种, 抗SCN3号生理小种, 产量表现较好, Hartwig抗SCN1、2、3、5、14号生理小种, 而且产量高[12]。晋1265高抗SCN1、3、4和5号生理小种, 且农艺性状优良[13]。中品03-5373是由晋1265与Hartwig杂交选育出的高抗SCN4号生理小种, 免疫14号生理小种[14], 经鉴定免疫SCN3的优良品系。
 | 图1 中品03-5373的系谱Unknow代表未知亲本。Unknow shows the parent unknown.Fig.1 Pedigree chart of Zhongpin 03-5373 |
1.2 分子标记的开发和筛选
1.2.1 InDel标记 根据灰皮支黑豆、中品03-5373及中黄13的重测序数据鉴定出的InDel位点[15], 在已报道的位于Gm18的控制SCN抗性主效QTL区间[16]及Gm12和Gm14共选取11个位点, 利用在线引物设计软件Primer3 (V.0.4.0) (http://frodo. wi.mit.edu/primer3/)设计引物, 引物由上海生工生物工程技术服务有限公司合成, 对设计的引物进行退火温度及PCR反应体系等条件的优化和筛选。通过6%变性聚丙烯酰胺凝胶电泳检测, 开发出11个条带单一的标记用于系谱材料基因型鉴定(表1)。
1.2.2 EST-SSR和SSR标记 用161个分子标记对11份系谱材料进行分子鉴定, 参照Song等[17]大豆公共遗传连锁图谱以及SoyBase数据库(http://www.soybase.org/)上公布的新SSR标记, 选择分布在20个染色体的150个标记(148个SSR标记和2个EST-SSR标记), SSR引物序列来自美国农业部大豆基因组数据库(SoyBase), EST-SSR引物序列来自Liu等[18], 结合开发出的11个InDels标记。上述引物均由上海生工生物工程技术服务有限公司合成。
| 表1 开发的InDel标记在染色体上的位置、引物信息及目标片段大小 Table 1 Chromosomal location, primer sequences and expected fragments size of nine InDel markers |
1.3 DNA提取和检测
从每份材料中取5粒种子种于温室育苗, 待长出三出复叶, 取其中一株的新鲜三出复叶, 在液氮中研磨, 依照快捷型植物基因组DNA提取系统(MBI Fermantas)提取参试材料的基因组DNA。用0.8%琼脂糖凝胶电泳检测提取DNA质量, 并用NanoDrop ND-1000 UV/VIS Spectrometer型紫外分光光度计(Thermo Fisher Scientific)检测DNA浓度, 将DNA稀释至10 ng µL-1后于4℃保存备用。
PCR反应体系20 µL, 包括45 ng基因组DNA、1×PCR缓冲液、0.2 mmol L-1 dNTPs、上下游引物各0.15 μmol L-1、1 U Taq聚合酶, PCR在PE9600上进行, SSR引物PCR反应条件为95℃ 5 min; 94℃ 40 s, 47℃ 40 s, 68℃ 40 s, 38个循环; 72℃ 5 min延伸, 于4℃保存。InDel引物PCR反应条件: 95℃ 5 min, 94℃ 40 s, 55℃ 40 s, 68℃ 40 s, 38个循环; 72℃延伸5 min, 于4℃保存。扩增产物经6%的聚丙烯酰胺凝胶检测多态性、印染显色、读板、室温干燥后统计条带照相。
1.4 数据调查和统计分析
选择条带清晰的标记进行统计, 根据扩增条带大小赋值, 建立原始数据群。用PowerMarker V. 3.25计算标记的等位变异数量(allele number)和基因遗传多样性(gene diversity), 以IBD方法寻找与大豆胞囊线虫抗性相关的候选标记, 用Microsoft Excel 2007分析。
系谱亲本与子代间的遗传关系采用共祖先系数、相似系数和遗传贡献率3种方法统计分析。共祖先系数法(coefficient of parentage, CP法)是一种理想状态下计算亲本对子代遗传贡献的方法, 双亲传递给子代2个等位基因中的一个, 故父母本对子代的遗传贡献率均为0.5。相似系数法(similarity coefficient, SC法), 不考虑每个标记在系谱中的传递情况, 只比较子代与亲本之间等位变异是否相同, 成对品种相似系数用Nei’s (1979), 计算公式为 Sij=2 Nij/( Ni + Nj)[19], Ni和 Nj分别表示材料 i和 j中的条带数目, Nij表示材料 i和 j共有的条带数目, 由Popgen V.1.32软件完成。遗传贡献率法(genetic contribution, GC法): 亲本A对后代X的贡献用 λA=>X, 亲本B对后代X的贡献用 λB=>X表示, A和X间的期望标记相似性为 SAX, 遗传贡献为: λA=>X=( SAX- SBX SAB)/1-( SAB)2[20]
在以上3种计算亲本对子代的遗传贡献率的基础上, 隔代亲本对子代的遗传贡献率是按照核遗传方式反推的方法计算祖先亲本对子代的遗传贡献。例如A是由B和C杂交形成, B和C对A遗传贡献率别为30%和70%, B是由D和E杂交形成, D和E对B的遗传贡献率分别为40%和60%, 那么祖先亲本D和E对A的遗传贡献率分别为12%和18%。
2 结果与分析
2.1 中品03-5373及其亲本的多态性分子标记 特点
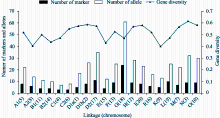
利用161个分子标记对11份材料鉴定, 有152个标记表现多态性, 包括SSR标记141个, EST-SSR标记2个和InDel标记9个, 每个标记的等位变异范围为2~5个, 平均为2.9个。这些多态性标记分布在20个染色体, 其中位于Gm18的标记最多, 为24个, 检测到等位变异61个; 以Gm04和Gm06染色体的标记最少, 为3个, 分别携带7个和8个等位变异(图2)。152个标记多态性信息指数(PIC)变化范围为0.1653~0.7769, 其中4个位点(Satt311, Satt152和Barcsoyssr_18_107)的多态性最高, 均为0.7769, 多态性最低的有6个位点(Satt178、Satt525、Satt561、Satt570、Satt573和Satt687), 均为0.1653, 平均为0.5224。各个染色体在基因多样性水平上存在差异, 以Gm03包含遗传多样性最高, 为0.6152, Gm09的遗传多样性最低, 为0.4 (图2)。在152个位点上检测到437个等位变异, 其中低频(<0.3)的等位变异比较多, 占等位变异总数的比例为54.7%; 发生频率范围为0.2~0.3的等位变异数最多, 为87个, 占等位变异总数的19.9%; 高频(> 0.9)等位变异数最少, 只有6个, 占总等位变异数的1.4% (图3)。4个SSR标记(Satt152、Satt179、Barcsoyssr_18_107和Satt196)形成的12种单倍型可以将11份材料区别开来(表2)。
 | 图2 152个位点在染色体上的分布和多样性Fig. 2 Distribution pattern and diversity of 152 loci on the chromosomes |
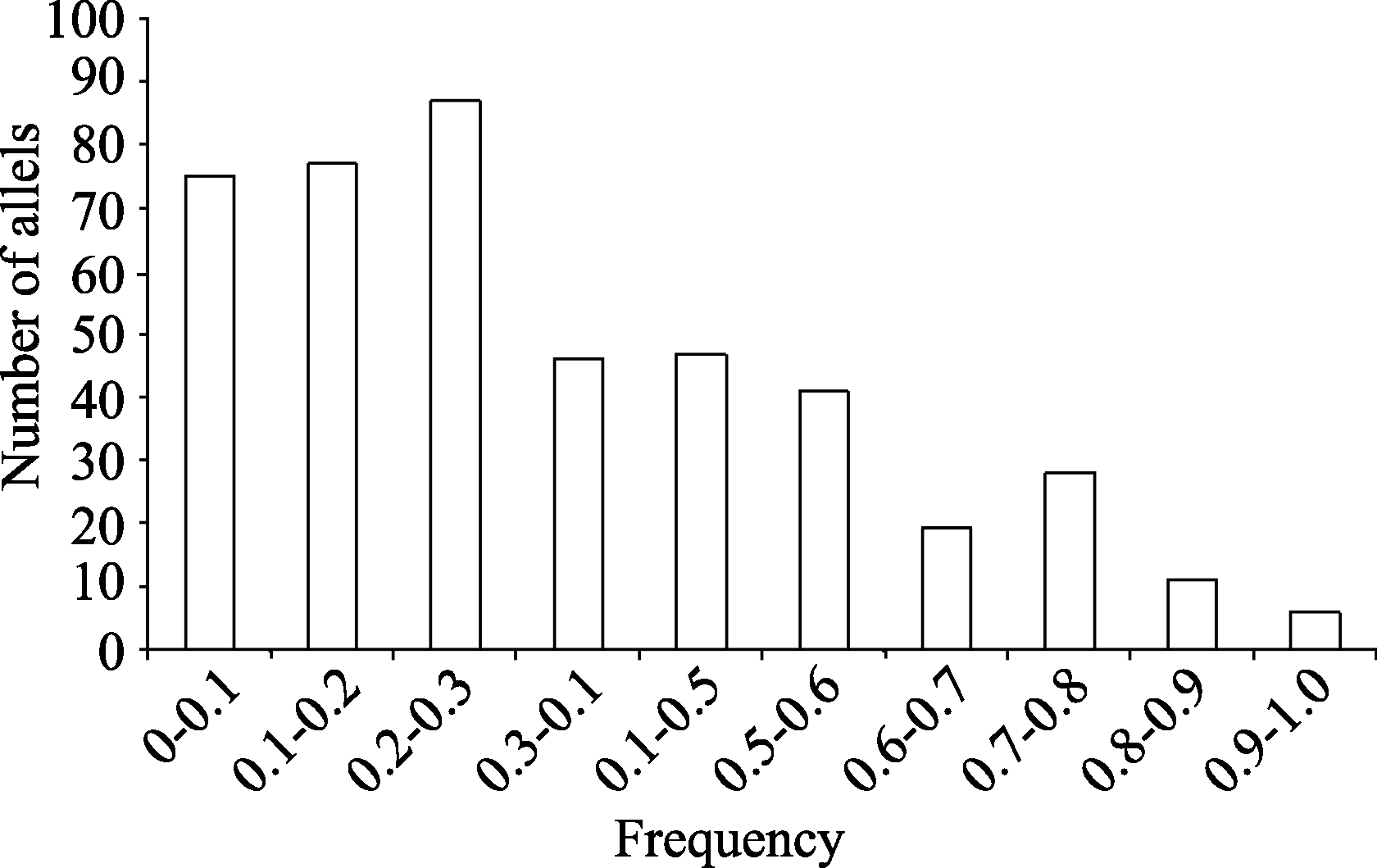 | 图3 437个等位变异发生频率分布Fig. 3 Frequency distribution of 437 alleles |
| 表2 4个SSR标记(Satt152、Satt179、Barcsoyssr_18_107和Satt196)形成品种特有单倍型 Table 2 Haplotypes formed by four SSR markers (Satt152, Satt179, Barcsoyssr_18_107, and Satt196) |
2.2 3种遗传贡献率方法的比较
CP法结果看出, 同一世代的亲本对中品03-5373的遗传贡献率是相同的, 离中品03-5373较近的亲本对其遗传贡献率要高于离其较远的亲本。PI437654、Forrest及灰皮支黑豆属于相同世代, GC法和SC法数据显示, Forrest对中品03-5373的遗传贡献率高于父本, 灰皮支黑豆是晋1265的母本, 灰皮支黑豆的遗传贡献率高于Forrest; Dyer和Bragg是同一世代, 2种方法结果都是Dyer的遗传贡献率高于Bragg。在Hill、Lee和Peking的组合中, Lee和Peking是一个世代, 要早于Hill所在的世代, 但在该组合中, Hill对中品03-5373的遗传贡献率在GC法中要低于Lee的遗传贡献率, 在SC法中低于Peking的遗传贡献率。祖先亲本对中品03-5373的遗传贡献率表现出母本高于父本的情况, 但在Hill、Lee和Peking的组合中出现例外, 即无论GC法还是SC法其结果都是晋1265对中品03-5373的遗传贡献率高于Hartwig, 表现出母本的遗传贡献率高于父本。SC法所揭示的遗传贡献率要显著高于GC法的( t测验, P<0.01), 虽然本实验2种方法揭示的亲本对子代的遗传贡献率的规律基本相同, 但GC法含有隔代传递的信息, 故揭示的遗传贡献率更接近真实水平(表3)。
相关性分析表明, 3种方法遗传规律基本相同, 相对于理想状态CP法而言, GC法和SC法之间的相关性更高一些( r = 0.943), GC法和CP法的相关性与SC法和CP法的接近, 相关系数分别为0.839和0.866。
2.3 SCN3抗性相关标记的追踪
中品03-5373系谱中不同组合间的重组率存在差异, 从双亲Dyer和Bragg到子代Forrest, 发生28次重组事件, 占双亲间多态性位点数(77)的35.9%, 在可追踪的3个组合中最高; 从双亲晋1265和Hartwig到子代中品03-5373发生的重组事件最多, 为33个, 占双亲间多态性位点数(107)的30.8%, 而Forrest和PI437654组合(子代为Hartwig)无论是重组事件(25个)还是重组事件发生频率(24.3%)在3个组合中都是最少的。
根据152个多态性分子标记等位变异分析, 与中品03-5373相同的等位变异且只存在抗病祖先亲本的分子标记有67个, 分布在除Gm01和Gm09之外的18个染色体上, 以Gm20分布所占比例最高(7/8)。其中7个抗病材料共有的等位变异位点有5个, 6个抗病材料共有的等位变异位点有1个, 5个抗病材料共有的等位变异位点有6个, 4个抗病材料共有的等位变异位点有8个。在这些抗病材料特有等位变异位点中只有49个位点可以追踪到抗病祖先亲本, 49个位点中有30个位点的等位变异由灰皮支黑豆经晋1265传递给中品03-5373, 这些位点中12个位点传递给中品03-5373的等位变异还存在于Peking及PI437654等抗病亲本中, 18个位点传递给中品03-5373的等位变异只存在于灰皮支黑豆、晋1265和中品03-5373, 在Gm18上存在8个这样的位点。能够确定是由PI437654将其等位变异传递给中品03-5373的位点有4个, 有3个(Satt311、Sat_365和Satt543)位点位于Gm17, 1个(Satt066)位于Gm14, 在Satt311和Satt543 (均在Gm17)位点, 由PI437654传递给中品03-5373的等位变异不存在系谱中的其他亲本, 在Sat_365和Satt066两位点上该等位变异还存在于Peking或灰皮支黑豆等抗性亲本中。其余15个位点可以追踪到灰皮支黑豆、Peking或PI437654这3个祖先抗性亲本, 但是这些等位变异的来源是模糊的(图4)。152个标记中还有部分标记能够追踪到抗性祖先亲本, 但是这些亲本传递给中品03-5373的等位变异也存在于感病亲本中。
9个位点可以将11份对SCN3反应不同的材料分辨开来(表4), 为大豆抗SCN3相关标记。对9个标记通过系谱追踪发现, 有3个标记(Satt309、Barcsoyssr_18_107和Satt723)按照方式一传递, 灰皮支黑豆通过晋1265传递给中品03-5373。位于Gm16 (LG J) Sct_193的等位变异同时为抗病品种PI437654、Hartwig、晋1265及灰皮支黑豆所拥有, 该等位变异来源模糊, 有可能按照PI437654、Hartwig传递到中品03-5373, 也有可能按照灰皮支黑豆、晋1265传递到中品03-5373, 传递到中品03-5373的等位变异既可能来自国外血缘也可能来自国内血缘(方式2)。另外5个标记(ESSR197、Satt197、Barcsoyssr_18_85、SCN_18_72和Sat_168)上发现8个抗病材料中特有的等位变异, 有可能来自系谱中的国内抗性亲本, 依照灰皮支黑豆, 晋1265传递给中品03-5373, 也有可能来自国外抗性血缘, 同时在国外抗性血缘的传递过程中存在模糊传递方式, 即Hartwig中的与大豆胞囊线虫3号生理小种抗性相关的等位变异既可能来自Peking, 也可能来自PI437654。
| 表3 各祖先亲本对中品03-5373的遗传贡献率 Table 3 Contribution rate of ancestor parents’ to Zhongpin 03-5373 |
 | 图4 152个标记在染色体(连锁群)上的分布及每个标记携带有与中品03-5373相同等变异的抗病材料在图谱右侧以圆表示抗病的品种。Circles indicate the cultivars with resistance.Fig. 4 Distribution of 152 markers on the chromosome (linkage groups) and the cultivars with the same allele as Zhongpin 03-5373 on the each marker |
| 表4 根据系谱追踪传给中品03-5373对大豆胞囊线虫3号生理小种抗性相关标记 Table 4 Markers associate with SCN3 based on the pedigree tracing |
3 讨论
系谱常用于研究材料间遗传关系, 但在实际应用中, 由于育种过程中偏亲选择(选择压)的存在, 目标基因的漂移, 未知亲缘亲本的存在都会导致研究者过高或者过低地估计品种间的遗传关系。近年来, 可以直接检测品种间遗传变异的分子标记已越来越多地被应用到验证已知系谱的正确与否、推测未知亲本及评估系谱材料间的亲缘关系[21,22,23]。Evans等[23]用全基因组的80个SSR标记检测307份苹果系谱材料, 结果显示, 11份栽培品种和4个品系的系谱中有一个亲本是错误的, 而文献报道的Dayton、Liberty和X-277的系谱皆是错误的。本研究中晋1265是抗大豆胞囊线虫病的重要抗源之一, 据记载是利用灰皮支黑豆与鲁豆4号杂交选育而成。本研究利用152个多态性分子标记鉴定结果显示, 晋1265与鲁豆4号的相似性很低, 在38个位点上携有非双亲等位变异, 占总标记数的29%, 可能本实验所用的鲁豆4号与选育时所用的亲本来源不同。有文献报道用于测序的Williams82, 其材料不同来源也存在差异[24]。
3.2 基于系谱抗SCN相关标记的鉴定
袁翠平等[25]分析我国已审定的80个抗(耐)病品种中, 25个有明确的系谱, 其中绝大多数(21个)品种的抗病基因来自Peking (美国从我国引进的北京小黑豆)或北京小黑豆, 2个品种来自哈尔滨小黑豆, 各有1个品种来自PI437654和灰皮支黑豆。抗病品种选育进展缓慢与胞囊线虫病表型性状鉴定难有关。因此, 培育抗SCN品种具有重要意义。
到目前为止, 已定位与大豆胞囊线虫抗性相关的QTL很多, Concibido等[16]对20年终抗大豆胞囊线虫QTL进行整理, 共收集60个与大豆抗胞囊线虫相关的QTL。由于所用的作图群体不同, 已定位到QTL局限到具体的某个群体[26]。本试验采用遗传基础比较广泛且包含国内外抗大豆胞囊线虫的优良抗源, 用系谱分析的方法寻找到9个与大豆胞囊线虫3号生理小种相关的候选标记。研究发现 rhg1控制着与SCN相关50%以上的变化, 同时对包括3号生理小种在内的多个生理小种都有抗性, 认为其为SCN抗性主效位点, Cregen等[27]发现Satt309和Sat_168是与 rhg1紧密连锁且共分离的, 本研究在Gm18上共发现5个抗性标记, Barcsoyssr_18_85、SCN- 18-72、Sat_168、Satt309和Barcsoyssr_18_107, rhg1位于Sat_168和Satt309之间, SCN-18-72和Barcsoyssr_18_107与 rhg1物理距离分别为18.7 kb和22 kb。Cook等[28]用基因沉默的方法验证 rhg1-b的3个基因作用控制所研究的抗性材料对大豆胞囊线虫的抗病性, rhg1-b位于用系谱分析方法寻找的抗性标记Barcsoyssr_18_85和SCN- 18-72之间。常玮等[29]对Concibido收集的60个与大豆抗胞囊线虫相关的QTL及SoyBase数据库(http://www.soybase.org/)上公布的96个抗性QTL筛选整理, 用图谱整合软件和元分析方法获得27个大豆抗SCN一致性QTL, Satt315-Sat_400是其获得的一个抗SCN3的QTL, 遗传贡献率为26.2%, 本研究在Gm08上发现一个抗性标记ESSR197, 位于该QTL区间, 表明利用系谱追踪抗性相关标记的方法是可行的。同时本研究鉴定的3个标记, 即Gm11上的Satt197, Gm16上的Sct_193和Gm19上的Satt723, 尚未见报到, 有可能是新的抗性位点。
4 结论
用161个分子标记鉴定高抗大豆胞囊线虫3号生理小种的优异品系中品03-5373的系谱发现, 在祖先亲本中以灰皮支黑豆贡献给中品03-5373的等位变异最多, PI437654次之。与材料间的抗病性相结合, 发现9个与胞囊线虫3号生理小种抗性相关的候选标记, 为抗大豆胞囊线虫病基因克隆和分子标记辅助抗病新品种选育提供了有益信息。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|